Mekanismer for biodiversitet
Oversatt herfra, {-kursiv og understreking ved oversetter.}
Nylig banebrytende vitenskapelig forskning som forklarer de virkelige mekanismene til biologisk mangfold:
Påstanden om at evolusjon er et faktum, gjentas som et mantra av tilhengere av evolusjon og naturalisme, som på den måten prøver å rettferdiggjøre sin vantro mot en intelligent skaper. En av de hyppigste påstandene er at mikroevolusjon og makroevolusjon er de samme på en annen tidsskala. Og at det ikke er noen mekanisme som hindrer mikro i å bli makro. De som er bedre informert, vet at mekanismen som provoserer frem endring og evolusjonære nyheter over artsnivået, det vil si endringen fra bakterier, som danner mennesket, er UKJENT.
For å forklare opprinnelsen til biologisk mangfold, kroppsform, kroppsplaner, organutvikling, og til slutt, hvis påstanden om makroevolusjon fra Luca til homo sapiens er sann, må vi først forstå hvordan organer, lemmer og skapninger oppstår, hvilken mekanisme bestemmer hvilken størrelse og form de skal ha når de vokser. Hvilken mekanisme programmerer cellen til å 'vite' hvordan og hvor den skal danne et organ? Hvordan vet skapningen hvordan man gjenoppbygger et avkuttet lem til riktig størrelse, form og orientering? Når dette er belyst, kan vi spørre om de samme mekanismene forklarer biologisk mangfold. Det ser ut til at banebrytende vitenskapelig forskning begynner å avdekke dette mangeårige mysteriet, og det er langt fra å bli forklart gjennom neodarwinistiske-spådommer og påstander, men epigenetiske mekanismer, som vil bli belyst nedenfor.
Bilde 1. ATP-motor med null-gradient
 Det er nå kjent mye om morfogen-gradienter i det utviklende embryoet - hvordan celler vet hvor de skal gå og hvilke typer celler de skal bli. Nyere forskning viser at elektriske potensialer i ikke-hjerneceller er et signal for å skape mønstre under utvikling og under gjenoppbygging av organer. Denne informasjonen om forskjellige feltpotensialer rundt individuelle celler kan gi informasjon for det utviklende organet. Nå viser forskning fra laboratoriet til Dr. Michael Levin nye måter som celler signaliserer med elektrisitet og den store betydningen av elektriske egenskaper for individuelle celler og vev. Han beskriver hvordan elektriske gradienter og felt er kritiske i 3D-funksjonen og formen til celler og organer.
Det er nå kjent mye om morfogen-gradienter i det utviklende embryoet - hvordan celler vet hvor de skal gå og hvilke typer celler de skal bli. Nyere forskning viser at elektriske potensialer i ikke-hjerneceller er et signal for å skape mønstre under utvikling og under gjenoppbygging av organer. Denne informasjonen om forskjellige feltpotensialer rundt individuelle celler kan gi informasjon for det utviklende organet. Nå viser forskning fra laboratoriet til Dr. Michael Levin nye måter som celler signaliserer med elektrisitet og den store betydningen av elektriske egenskaper for individuelle celler og vev. Han beskriver hvordan elektriske gradienter og felt er kritiske i 3D-funksjonen og formen til celler og organer.
Et fremtredende aspekt ved flercellede skapninger er at de har organer av en bestemt størrelse og form. Når regenerering skjer hos reptiler, vokser den samme nøyaktige formen ut igjen. Informasjon for den cellulære aktiviteten ser ut til å eksistere i rommet som vil utgjøre det spesifikt formede organet. Levin bemerker at "kreft kan sees på som en geometrifeil, fordi tumorceller vokser, migrerer og fungerer uten hensyn til den ordnede strukturen de oppstår innenfor."
Hver celle har spesifikke elektriske gradienter og egenskaper som til sammen danner et stort elektrisk informasjonsfelt. Dette informasjonsfeltet kan vise individuelle celler i embryoet hvordan de skal oppføre seg. Dette er analogt med det faktum at den elektriske flyten mellom celler i det tidlige embryoet, danner det grunnleggende nettverket som deretter bygges inn i en formell struktur med forseggjorte kjemiske synapser. Dette skjer også under gjenoppbygging av vev. På en eller annen måte styrer informasjonen den elektriske strømmen gjennom synapsene for elektriske veikryss i den fremtidige strukturen. Elektriske signaler ser nå ut til å være kritiske i formen av organer, selve funksjonen og identiteten til organer, og skapelsen av nye lemmer på dyr som regenererer. Hos disse dyrene styres stamcelle-adferd av strømmer skapt med kalium, natrium, klorid og protoner, som påvirker de genetiske nettverkene til celler på avstand. Nyere forskning viser at lemmer kan påvirkes av proton- og natriumendringer. Nye typer ionekanaler, pumper og elektriske forbindelser er nå funnet i en rekke forskjellige organer.

Bilde 2: Mekanisme for biodiversitet
Dette er bemerkelsesverdig. Hvis elektriske veikryss-synapser bestemmer celleform, bør makroendringer, som vanligvis tilskrives makroevolusjon, og strukturelle biologiske nyheter også avhenge av disse.
Elektrisk signalering er nøkkelen for at cellene skal kunne tolke miljøet sitt på riktig måte, og når denne prosessen går galt, går cellene som standard til et kreftprogram. (4)
Mens ionestrømmer kontrollerer atferd på cellenivå som migrasjon, differensiering og spredning, fungerer bioelektriske signaler også som hovedregulatorer av storskala form i mange sammenhenger: Et enkelt signal kan indusere komplekse, svært orkestrerte, selvbegrensende, nedstrøms -morfogenetiske kaskader. For eksempel kan en umodulert fluks av protoner forårsake dannelse av en komplett hale av riktig høyde og vevs-sammensetning.
Våre data tyder på at mekanismen som blastema-celler bruker til å spørre resten av verten (for å bestemme hvor såret er lokalisert og hvilket annet vev som allerede finnes i fragmentet og dermed ikke trenger å gjenskapes) er mediert av fysiologiske signaler som passerer gjennom nerver og knutepunkt-baner på (lang)distanse.
En betydelig komponent av morfogenetiske signaler er ioniske av natur. Bemerkelsesverdig nok er imidlertid denne effekten ikke-lokal av natur - det er transmembran-potensialet til andre, ganske fjerne celler som bestemmer den metastase-lignende effekten (eks. ved spredning av kreft).
Bilde 3. Bioelektrisk felt styrer utvikling til frosk
 Mange kritiske spørsmål gjenstår om hvordan cellulær polaritet synkroniseres og forsterkes på tvers av embryonale felt, for å tillate celler å fastslå sin posisjon i forhold til midtlinjen. Vi identifiserte en avhengighet av asymmetrisk genuttrykk i tidlig kommunikasjon mellom venstre og høyre side hos kyllingen og frosken. For eksempel avhenger uttrykket av venstresidige markører av hendelser som skjer på høyre side, i veldig tidlige stadier, noe som antyder at de to sidene må koordinere sin beslutning med hensyn til L-R-identiteten til hver. En mekanisme for å kommunisere mellom celler og vev involverer veikryss: multimerer av connexin-proteiner danner kanaler mellom celler og passerer små molekyler, underlagt kompleks regulering av forskjellige signaler.
Mange kritiske spørsmål gjenstår om hvordan cellulær polaritet synkroniseres og forsterkes på tvers av embryonale felt, for å tillate celler å fastslå sin posisjon i forhold til midtlinjen. Vi identifiserte en avhengighet av asymmetrisk genuttrykk i tidlig kommunikasjon mellom venstre og høyre side hos kyllingen og frosken. For eksempel avhenger uttrykket av venstresidige markører av hendelser som skjer på høyre side, i veldig tidlige stadier, noe som antyder at de to sidene må koordinere sin beslutning med hensyn til L-R-identiteten til hver. En mekanisme for å kommunisere mellom celler og vev involverer veikryss: multimerer av connexin-proteiner danner kanaler mellom celler og passerer små molekyler, underlagt kompleks regulering av forskjellige signaler.
Så det virker som om det er innenfor elektriske veikryss, at handling skjer, men handlingen i seg selv er REGULERING OG KOMMUNIKASJON GJENNOM ULIKE SIGNALER. DET ER NØKKELEN.
Vi viste at elektriske veikryss er avgjørende involvert i L-R-mønste i tidlige embryoer av Xenopus og kylling. Veikryss er et bioelektrisk mønsterelement som setter opp domener av isopotensielle cellefelt under morfogenesen.
Serotonin-signalering brukes i informasjonsutveksling mellom celler i prosesser som L-R-mønster og kontroll av timing og cellebevegelse under gastrulering. Vi har vist at serotonin brukes av både kylling- og froskeembryoer, i svært tidlige stadier, som et lite molekylsignal som transporteres i en venstre-høyre gradient og regulerer utviklingen av lateralitet. Faktisk vet vi nå at det tidlige froskeembryoet bokstavelig talt er et elektroforese-kammer, som bruker spenningspotensialer til å generere konsekvente partiske venstre-høyre gradienter i serotonin i en epigenetisk prosess som ikke er avhengig av zygotisk genuttrykk. Vi har modellert denne prosessen kvantitativt, og karakterisert nye intracellulære serotoninbindende proteiner som direkte aktiverer asymmetriske genuttrykk etter deres bevegelse til høyre, og kobler en tidlig biofysisk prosess til transkripsjons-regulering via kromatin-modifikasjonsveier. Serotonin er også en nøkkel-mediator for bioelektrisk kontroll av nevronal utvekst fra transplantasjoner.
Elektriske felt styrer 3D-form av celler og organer
Bilde 4. Bioelektrisk kode i nevron
 Hvordan vet cellen hvilken størrelse og form den skal ha? Mange celler endrer form for å gi forskjellige funksjoner, som mikroglia. Enda mer komplekst er spørsmålet om hvordan organer, lemmer og skapninger vet hvilken størrelse og form de skal ha når de vokser. Hvordan vet cellene hvordan og hvor de skal danne et organ? Hvordan vet skapningen hvordan man gjenoppbygger et avkuttet lem til riktig størrelse, form og orientering?
Hvordan vet cellen hvilken størrelse og form den skal ha? Mange celler endrer form for å gi forskjellige funksjoner, som mikroglia. Enda mer komplekst er spørsmålet om hvordan organer, lemmer og skapninger vet hvilken størrelse og form de skal ha når de vokser. Hvordan vet cellene hvordan og hvor de skal danne et organ? Hvordan vet skapningen hvordan man gjenoppbygger et avkuttet lem til riktig størrelse, form og orientering?
Det er tusenvis av de samme spørsmålene, inkludert hvordan astrocytter og nevroner vet de nøyaktige nettverkene de skal danne. Hvordan vet immunceller hvordan de skal reise gjennom svært komplekse forskjellige 3D-miljøer? Et like vanskelig spørsmål har blitt tatt opp i tidligere innlegg om hvordan cellen vet nøyaktig hvilken form en kodet sekvens i et protein vil ha. Både celler og mikrober endrer kodene til deres proteintoksiner, som krever ekstremt detaljerte og nøyaktige former. Faktisk kan moderne vitenskap ikke beregne hvilken form en 400 aminosyrekodet sekvens vil ha når den foldes til et protein. Det ville ta alle superdatamaskinene til sammen to tusen år å beregne foldingen av ett gjennomsnittlig protein. Likevel samles proteiner til den nøyaktige formen på et millisekund, hjulpet av svært komplekse chaperone-molekyler. Celler redigerer rutinemessig budbringer-RNA med alternativ spleising for å lage en hel rekke former. Hvordan vet de hvordan de skal gjøre dette?
Forskning viser at: Hvis en celle fra et tocellet embryo fjernes, blir den fortsatt hele skapningen. Utsultede flatormer krymper og holder perfekte proporsjoner mellom organene.
Planaria kan reprodusere hele kroppen fra et lite stykke.
Amfibier kan vokse ut et perfekt proporsjonert lem.
Mange strukturer trenger nervemediert informasjon for å opprettholde form og funksjon.
For alle disse spørsmålene er det vanskelig å forestille seg at informasjonen om 3D-formen er inne i én celle, eller til og med en gruppe celler. Det virker mer sannsynlig at et informasjonsfelt på en eller annen måte styrer disse prosessene. Er dette beviset for et elektrisk felt med informasjon som danner 3D-former av celler, organer og skapninger, også relatert til sinnets påvirkning?
Elektriske gradienter og felt i og rundt celler
Tidligere innlegg (fra denne forfatteren) har vist den kritiske delen som elektriske synapser spiller for å danne strukturen til det nevronale nettverket, som bruker kjemiske synapser. Et tidligere innlegg viste også at feltpotensialer i og rundt hjerneceller, selv om de er dårlig forstått, er viktige for spesifikke funksjoner. Flere innlegg har dokumentert den forseggjorte kommunikasjonen som skjer mellom celler gjennom signalering, inkludert nevrotransmittere, hjernefaktorer, cytokiner og hormoner.
En betydelig mengde forskning viser betydningen av ulike gradienter i rommet mellom cellene for kommunikasjon. Dette ble notert i innleggene  om bevegelse av blodplater som tiltrekker og signaliserer til andre immunceller. Et annet innlegg viste de forseggjorte reisene til leukocytter. Det er nå kjent mye om morfogen-gradienter i det utviklende embryoet - hvordan celler vet hvor de skal gå og hvilke typer celler de skal bli.
om bevegelse av blodplater som tiltrekker og signaliserer til andre immunceller. Et annet innlegg viste de forseggjorte reisene til leukocytter. Det er nå kjent mye om morfogen-gradienter i det utviklende embryoet - hvordan celler vet hvor de skal gå og hvilke typer celler de skal bli.
Bilde 5. Ioner rundt membran
Nyere forskning viser at elektriske potensialer i ikke-hjerneceller er et signal for å skape mønstre under utvikling og under ombygging av organer. Denne informasjonen om forskjellige feltpotensialer rundt individuelle celler kan gi informasjon for det utviklende organet. Nå viser forskning fra laboratoriet til Dr. Michael Levin nye måter som celler signaliserer med elektrisitet og den store betydningen av elektriske egenskaper for individuelle celler og vev. Han beskriver hvordan elektriske gradienter og felt er kritiske i 3D-funksjonen og formen til celler og organer. Professor Levin bemerker at kreftceller bruker elektriske gradienter til mange formål. Faktisk kan avvik i elektriske prosesser føre til kreft. Forskjeller i elektrisk potensial i celler er viktige for å avgjøre om en kreft metastaserer og elektriske tilstander i en celle kan ha effekter på fjerne celler, og utløse metastaser. Disse elektriske potensialene i en celle kan signalisere andre celler som utløser genetiske nettverk og epigenetiske effekter.
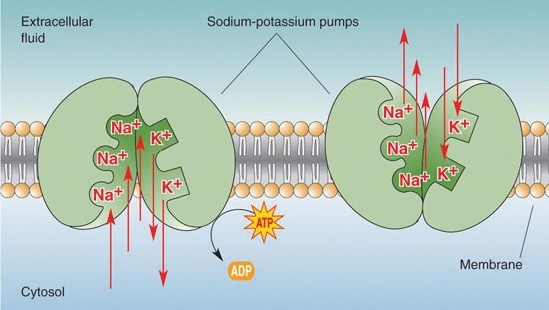
Den elektriske tilstanden til en celle bestemmes av mange komplekse faktorer. Den ene er åpning og lukking av ionekanal-proteiner, som kontrollerer bevegelsen av ladede partikler (ioner) over cellemembranen. Endringer i cellenes elektriske potensial via aktiviteten til ionekanaler har vist seg å kunne undertrykke eller utløse kreft. En annen faktor er elektriske synapser som overfører elektrisitet fra en celle til en annen.
Ulike elektriske egenskaper til celler påvirker hvilken type celler som er laget av stamceller, hvor mye reproduksjon som foregår, hvor cellene reiser til, og deres form og orientering. Elektriske gradienter og felt påvirker sikkert reisende immunceller, stamceller og alle hjerneceller. De utøver også dype effekter på andre somatiske celletyper under embryonal utvikling og regenerativ reparasjon. Elektriske egenskaper er samtidig med kjemisk metabolisme i bruk av genetisk nettverksbehandling. De er avgjørende for å helbrede sår og reagere på infeksjoner. De bestemmer symmetrien og formen til organer.
Bilde 6. Pumpe driver ioner gjennom cellemembranen
 Differensiell ionekanalaktivitet i kroppen setter opp spesifikke mønstre av spenningspotensial gjennom vev. Under utvikling bestemmer disse elektriske egenskapene til cellene hvilke typer celler de blir. For eksempel har to spesifikke grupper av celler i embryoet økt elektrisk potensial og disse blir de to øynene. Forskning viser at ved å endre det elektriske potensialet til celler på et kritisk tidspunkt når øynene lages, kan øyne induseres til å dannes andre steder, og effektivt omprogrammere andre celletyper til et komplett visuelt organ.
Differensiell ionekanalaktivitet i kroppen setter opp spesifikke mønstre av spenningspotensial gjennom vev. Under utvikling bestemmer disse elektriske egenskapene til cellene hvilke typer celler de blir. For eksempel har to spesifikke grupper av celler i embryoet økt elektrisk potensial og disse blir de to øynene. Forskning viser at ved å endre det elektriske potensialet til celler på et kritisk tidspunkt når øynene lages, kan øyne induseres til å dannes andre steder, og effektivt omprogrammere andre celletyper til et komplett visuelt organ.
Mens elektrisk aktivitet kan drive nedstrøms genetiske endringer, kan bare endring av den elektriske gradienten endre cellens oppførsel dramatisk. Etter en amputasjon forårsaker kontroll av spenningspotensialene til sårceller gjenvekst av et lem. Spesifikasjonen av detaljene ser ut til å eksistere i det elektriske feltet. I spesifikke områder av dyrets kropp vokser enten en hale eller lem på grunn av den elektriske signaleringen, som kickstarter et "bygg hva som kan passere"-program.
Elektriske felt som blir oppdaget med flere påvirkninger kan inneholde denne 3D-informasjonen. Endring av det elektriske feltet endrer informasjonen og de biologiske resultatene.
Oversatt versjon av artikkel fortsetter her.
Lenker:
Oversettelse og bilder ved Asbjørn E. Lund